
m6A-RNA甲基化阅读器YTHDF作用新模型
 2020-07-15
2020-07-15 
 MCE小分子
MCE小分子
N6-甲基腺苷 (m6A) 是哺乳动物 mRNA 上最常见的转录组修饰,m6A 修饰也是近年来新兴的研究领域。近期,顶级期刊 Cell 上就刊登了一项关于 mRNA 甲基化的最新研究,快跟小编一起来看科学前沿吧!
近期,美国康奈尔大学 Samie R. Jaffrey 研究组在 Cell 上发表了题为 “A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA” 的研究,揭示了 YTHDF 蛋白调节 m6A 修饰的 mRNA 的功能统一模型。
与“不同的 m6A 位点结合不同的 DF 蛋白”的主流观点不同,该研究人员发现,所有 m6A 位点与三个 DF 蛋白都以基本相似的方式结合,它们以冗余的作用方式诱导同一子集的 mRNA 降解,没有证据表明它们能直接促进翻译。

图 1. YTHDF 蛋白调控 m6A 修饰的 mRNA 的功能模型
m6A (N6-methyladenosine):是指腺嘌呤核苷 N6 位置发生了甲基化修饰。m6A 是真核 mRNA 最普遍的内部修饰,在功能上调节真核转录组,影响 mRNA 的剪接、输出、定位、翻译和稳定性。
m6A 修饰有三类调节器:
Writer 甲基转移酶 (MTC),负责催化,例如 METTL3、METTL14 和 WTAP;
Eraser 去甲基化酶 (Demethylase),负责去除甲基化,例如 FTO 和 ALKBH5;
Reader 直接识别和结合 m6A 位点,使 m6A 修饰的 RNA 发挥特定的作用,主要包括 YTHDC1、YTHDF1、YTHDF2、YTHDF3 等。
YTHDF:一种 m6A “阅读器”,即 Reader。胞质中的 YTHDF 家族包括了 YTHDF1、YTHDF2 和 YTHDF3 三种旁系同源物 (paralogs),也可以简称为 DF1、DF2、DF3。据报道,三种 DF 有不同的功能,DF1 促进 mRNA 的翻译,DF2 促进 mRNA 的降解,DF3 促进翻译和降解。
关于 DF 旁系同源物选择性地结合不同的 m6A 位点的机制目前尚未清楚。此外,DF 蛋白功能差异的机制基础也仍不清楚,尤其是在它们同源性很高的情况下。

图 2. 研究亮点
首先,研究人员观察 DF1 和 DF2 YTH 结构域与含有 m6A 的 RNA 的结合,发现在整个转录组中,DF 蛋白结合 m6A 的氨基酸以及结合近端的氨基酸都是保守的,因此三个 DF 蛋白与 m6A 结合的结构机制是相同的。
DF 旁系同源物结合 m6A 序列基序 (Sequence motifs) 的差异反映可以其结合特性,研究发现 m6A 残基几乎只存在 DR-m6A-CH (D = A, G, U; R = A, G; H = A, C, U) 这一个高度保守的序列基序中,像 GG-m6A-CU、AG-m6A-CU 都属于 DR-m6A-CH 的亚基序 (Submotif) 。
为了确定每种 DF 的结合偏好,研究人员通过全转录组 iCLIP 图谱,检测了 HEK293T 细胞内源性表达的 DF1、DF2 和 DF3 的结合位点,分析发现三种 DF 结合位点序列亚基序的偏好相同。再结合已报道的 PAR-CLIP 数据,发现即使用不同的细胞系和不同的 CLIP 方法,DF 旁系同源物的 RNA 结合基本相同。因此, DF 旁系同源物的 m6A 结合模式相同。
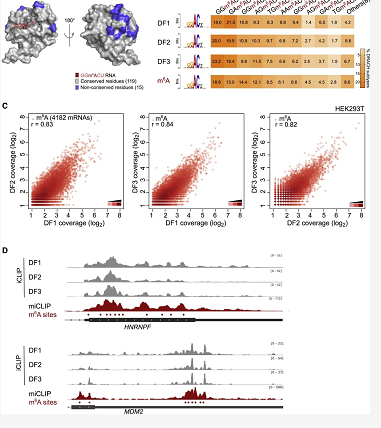
图 3. DF 蛋白与 m6A 结合位点的转录组分析
但是,不同的 DF 如何对 m6A-mRNAs 发挥不同的分子效应这个问题仍未解决,研究人员又分析了 DF 旁系同源物之间的效应区域差异,以及它们的相互作用蛋白 (不同的 DF 蛋白可能通过与不同的蛋白相互作用介导其不同的功能)。结果显示,DF 旁系同源物的序列、功能域、相互作用蛋白和细胞内定位都高度相似。
此外,研究人员还发现 DF 蛋白与降解有关的因子相互作用可信度较高,与翻译相关的因子相互作用可信度较低。
根据已有报道,研究人员考虑到每种 DF 蛋白都有介导 m6A-mRNA 降解的可能性。于是,他们利用 siRNA 选择性敲降 HeLa 细胞中的 DF1、DF2 和 DF3,使用 RNA-seq 检测 mRNA 的丰度,证实是 DF2 影响了 m6A-mRNA 稳定性,而非 DF1 或 DF3。

图 4. DF 蛋白冗余地控制 m6A-mRNA 的丰度和稳定性
再对 DF 进行不同组合的双重敲降和三重敲降,并利用放线菌素 D (Actinomycin D) 抑制转录后 m6A-mRNA 的水平证明了:DF 蛋白的联合活性导致 m6A 修饰的 mRNA 降解,DF 旁系同源物在功能上可以互相补偿,这种补偿功能也受其表达水平限制。当三种 DF 都耗竭时,补偿就不会发生,这时 m6A-mRNA 的稳定性得到最大程度的提高。
接下来,研究人员检测了 DF 在 m6A-mRNA 的翻译调控中的作用。三种 DF 都与多聚核糖体组分无关,而富集于信使核糖核蛋白 (mRNP) 组分,该结果与 “DF 蛋白(任何一种) 稳定地结合至 mRNA 3'UTR,增强其翻译的模型” 不一致。
任何的 DF 旁系同源物基因沉默都不影响 HeLa 细胞中 mRNA 的翻译效率,即使是 DF 三重敲降也没有显著降低翻译效率。因此,任何一种 DF 同源旁系物都不能直接增强 mRNA 的翻译,相反的,它们的主要作用是介导 m6A-mRNA 降解。
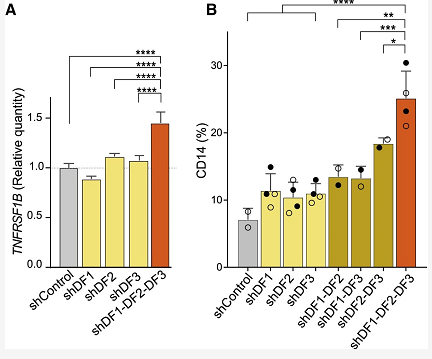
图 5. DF 蛋白冗余地抑制白血病细胞的分化
已有报道 DF2 在急性髓系白血病 (AML) 中过表达,与 AML 的发生和发展有关,DF2 的缺失导致一些抑制 AML 相关基因转录本上调,例如在 DF2 敲除的白血病前期细胞中,TNFRSF1B 转录本的半衰期增加,表面 TNFR2 的表达量上升。
为探索 TNFRSF1B 的抑制是否是通过三种 DF 蛋白共同作用介导,研究人员对 MOLM-13 白血病细胞中的 DF 进行单独或联合敲降,发现 TNFRSF1B mRNA 表达水平受 DF 的单独敲降影响较小,其中仅 DF2 的敲降使其略微升高,在三重敲降后却显著上调。因此,在 MOLM-13 细胞中 DF 蛋白共同作用控制 m6A-mRNA 的表达水平。

