
关于CRISPR系统的讲解
 2020-06-05
2020-06-05 
 MCE小分子
MCE小分子
很久很久以前,细菌就一直在与病毒或入侵的核酸进行斗争,为了在这场战疫中存活下来,细菌部署了多种防御机制,包括基于遗传因素的自适应型免疫系统,即 CRISPR。
在许多细菌和古细菌中, CRISPR 形成了独特的遗传基因座,这些基因座特异性靶向病毒核酸,从而获得了针对病毒的免疫力。且随着时间的推移建立起可遗传的编码免疫力。与此同时,病毒也不断在改变逃避策略,来绕开 CRISPR 系统。
于是,在生命进化史上,CRISPR 系统便成了细菌和病毒进行斗争产生的免疫武器。
CRISPR/Cas
CRISPR/Cas 系统——由一小段 RNA 和一种高效的 DNA 切割酶 (Cas 核酸酶) 组成的系统。
CRISPR (Clustered regularly interspacedshort palindromic repeats; 成簇的规律间隔的短回文重复序列) 是一种抵御外源 DNA 的免疫机制。
CRISPR 主体为 CRISPR 基因座 (CRISPR Array): 由多个重复序列 (Repeats)和间隔序列 (Spacers) 组成。
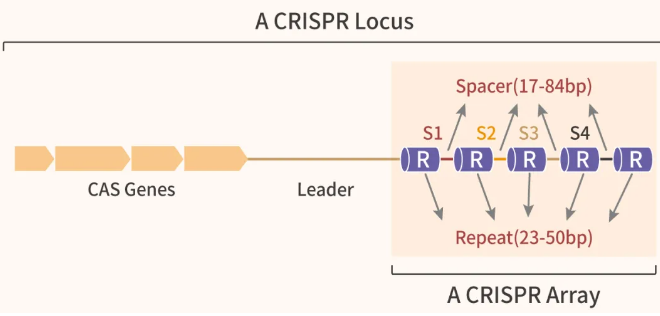
图 1. CRISPR/Cas 结构[2]
Cas (CRISPR-associated genes; CRISPR 相关基因): 可在 CRISPR 序列左侧 (图 1) 或右侧。
Leader (前导序列): 300-500 个碱基组成,位于 CRISPR 的 5' 端,与第一个重复序列直接相连,是一段相对保守的 AT 富集区。Leader 是新插入序列的识别位点,也能作为启动子,启动 CRISPR 基因座的转录。有些细菌无前导序列,如大肠杆菌。
CRISPR/Cas9
在细菌及古细菌中,CRISPR 系统共分成 3 类,其中 I 类和 Ⅲ 类需要多种 CRISPR 相关蛋白 (Cas 蛋白) 共同发挥作用。而来自 Streptococcus pyogenes 的 Ⅱ 型系统只需要一种 Cas 蛋白 (Cas9) 即可发挥核酸内切酶活性。因此,CRISPR/Cas9 系统应用最为广泛。CRISPR/Cas9 系统已经成功应用于植物、细菌、酵母、鱼类及哺乳动物细胞。
CRISPR/Cas9 作用机制
第一步: 捕获外源 DNA
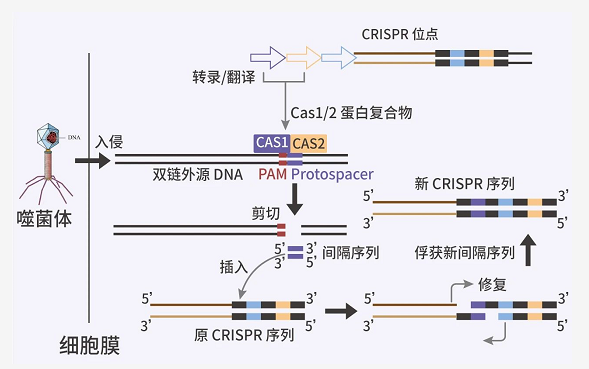
图 2. CRISPR/Cas9 操作第一步[6]
当噬菌体等外源 DNA 入侵时,CAS1/2 复合体通常识别 3' 端含 NGG 的前间隔序列邻近基序 (PAM) 区域。然后,前间隔序列 (Protospacer) 被剪切下来,作为间隔序列插入宿主 CRISPR 位点中,由重复序列隔开。PAM 序列结构简单,几乎可在所有基因中找到,因此得到广泛的应用。
第二步: crRNA 合成

图 3. CRISPR/Cas9 操作第二步[6]
CRISPR 序列转录,形成前体 CRISPR RNA (pre-crRNA),然后在 Cas9 蛋白的帮助下,被剪切成成熟的 tracrRNA-crRNA 双链 RNA 。II 型 CRISPR/Cas9 系统中,这一步由 Cas9 及 RNase III 共同完成。对 pre-crRNA 的剪切需要先合成一段反式激活 crRNA (tracrRNA)。tracrRNA 中有部分碱基序列与 pre-crRNA 上的重复序列存在严格的碱基互补关系。Cas9 及 RNase III 只有在 tracrRNA 与 pre-crRNA 上的重复序列配对形成双链 RNA 的条件下,才能对 pre-crRNA 进行剪切,剪切后,形成成熟的 tracrRNA-crRNA 双链 RNA。tracrRNA-crRNA 双链体被称为 gRNA。Cas9 核酸酶和 gRNA 形成 Cas9 核糖核蛋白 (RNP)。
改造后的 CRISPR/Cas9 将 tracrRNA 和 crRNA 融合成一条单链引导 RNA (sgRNA)。sgRNA 可以从单个质粒表达,用于直接转染或包装成用于病毒转导的颗粒。
另外,可通过与一些荧光物质融合,来可视化活细胞中的内源性基因组位点。如下图的荧光小分子 DFHBI (HY-110250)。
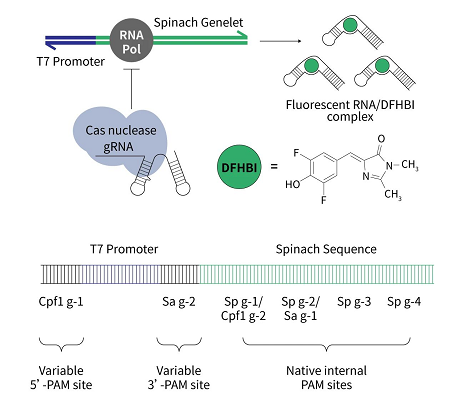
图 4. DFHBI 用于检测 Cas 核酸酶活性[8]
第三步: 靶向干扰

图 5. CRISPR/Cas9 操作第三步[6]
当外源 DNA 再次进入细胞时,Cas9 蛋白携带 gRNA,去识别外源 DNA 的 Protospacer (前间隔序列),并与之结合,通过 Cas9 解旋酶和核酸酶对靶基因进行剪切。造成靶基因 DNA 的双链断裂 (DSB),从而达到干扰靶基因表达的目的。
Cas9 的酶切活性被激活的两个条件:
1. 通过 crRNA 中的间隔序列与靶基因 DNA 的 Protospacer 互补配对。
2. Cas9 蛋白必须识别靶基因 DNA 上的 PAM 序列 NGG。
综上,CRISPR 技术主要是利用位点特异 Cas 核酸酶在基因组靶位点处引入 DNA DSB,再经细胞自身的非同源末端连接 (NHEJ) 或同源重组修复 (HDR) 对 DSB 进行修复,最终实现目标基因敲除和碱基编辑等基因组遗传修饰。
DNA 修复
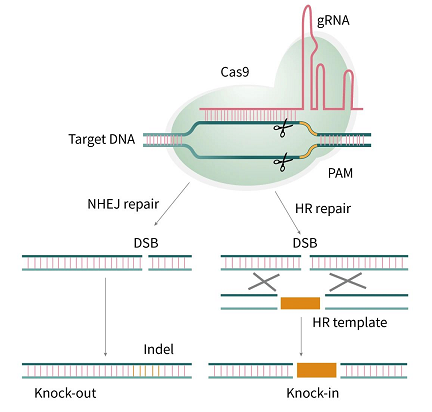
图 6. DNA 两种修复机制[6]
非同源末端连接 (NHEJ): 哺乳动物中很普遍,是 DNA 双链断裂的主要修复方式,多种修复酶强行将两个 DNA 双链断端连接一起,是一种易错修复,易在 DSB 位点出现 DNA 碱基的缺失或者插入,引起基因编码区阅读框的改变。NHEJ 修复过程不依赖于同源 DNA 序列,核心成分是具有催化作用的 DNA-PK,Ku70 和 Ku80,及 DNA 连接酶 IV 等。NHEJ 不限于细胞周期的某个阶段,可在细胞周期的任一时期发生。
同源重组修复 (HDR): HDR 修复过程需要姐妹染色单体作为修复模板,以同源重组的方式在 DSB 处生成所需要的序列替换,实现基因定点删除,插入,或者基因矫正,具有准确性和复杂性。而 HDR 在细胞周期的 S 和 G2 阶段起作用。
在细胞内,NHEJ 的发生率远高于 HDR,为了提高同源重组介导的精准修复就需要提高 HDR 的发生频率。通过抑制 NHEJ 修复途径的发生可以提高同源重组修复的效率。
CRISPR/Cas9 介导的 NHEJ 与 HDR
CRISPR 系统靶向结合特异性的 DNA 序列,诱导靶 DNA 双链发生精确切割。DSB 的产生刺激 DNA 损伤修复过程。提高基因组精准编辑效率。
抑制 NHEJ 修复途径相关化合物
SCR7
是有效的 DNA 连接酶 IV 抑制剂,DNA 连接酶 IV 是 NHEJ 途径中的关键酶。
高度选择性的 DNA-PK 抑制剂,DNA-PK 在 NHEJ 修复中起关键作用。
增强 HDR 修复途径相关化合物
RS-1
HDR 激活剂,可提高 Cas9 介导的敲入效率。
Nocodazole
细胞周期抑制剂,将细胞周期阻滞在 G1/S 期或者 G2/M 期,以提高 HDR 效率 (NHEJ 修复途径可在细胞周期的任一时期发生,而同源重组修复途径只在细胞周期的 S 期以及 G2 期发生)。
β3-肾上腺素能受体激动剂; 增强 CRISPR 介导的 HDR 效率。
蛋白转运抑制剂,抑制蛋白从内质网向高尔基体转运; 增强 CRISPR 介导的 HDR 效率。
CRISPR 的应用
CRISPR 技术已经用于多种不同疾病的研究,包括遗传性血液病镰刀状细胞贫血症,莱伯氏先天性黑蒙症,泰伊-萨克斯二氏病等。基于 CRISPR/Cas 系统而来的高通量基因组编辑技术和低成本突变检测方法也在禾谷类作物中不断得到发展与应用。
莱伯氏先天性黑蒙症 (Leber congenitalamaurosis; LCA) 是一种由中心体蛋白 290 (CEP290) 基因突变引起的遗传性视网膜变性疾病,由至少 18 个不同基因的突变引起。该病是遗传性儿童失明最常见的原因。眼科基因编辑疗法 AGN-151587 (EDIT-101) 已进入 I/II 期临床研究 NCT03872479。
AGN-151587 是一种腺相关病毒 AAV5 病毒载体,装载了 3 个成分,包括: 由 U6 聚合酶 III 启动子控制的 2 个指导 RNA (gRNA) RNA-323 和 gRNA-64,以及通过光受体特异性 GRK1 启动子表达的金黄色葡萄球菌 Cas9。
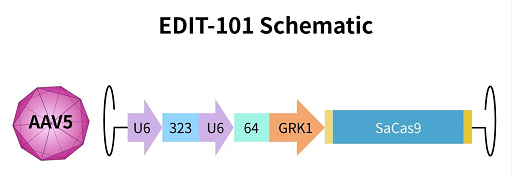
图 7. EDIT-101 示意图[9]
不同剂量的 AGN-151587 注射到视网膜下,将基因编辑系统递送到感光细胞中。当感光细胞表达基因编辑系统时,gRNA 指导的基因编辑可以消除或逆转 CEP290 基因上致病的 IVS26 突变,从而改善感光细胞功能。由于 Cas9 由光受体特异性 GRK1 启动子控制,因此该系统只在感光细胞内发生基因编辑,从而将潜在副作用最小化。CRISPR 基因编辑是永久性的,意味着可能只需要一次注入就可以终身改善视力。
CRISPR gRNA/Cas 9 质粒载体
SpCas9 载体
来源于化脓性链球菌,是一种高效、精准的基因编辑载体,编辑范围大,但是不能被包装在 AAV 病毒中用于基因治疗。
当与 gRNA 序列结合后,这些 Cas9 蛋白可在特定位点上进行基因组双链断裂。因其简捷性,特异性以及高效性,在基因组编辑中广泛使用。SpCas9 出现两个连续的 G 序列就可以进行编辑,SaCas9 需要四个碱基组合才能编辑,这样出现的频率低。
SaCas9 载体
来源于金黄色葡萄球菌,是第一个被用于基因治疗研究的小型 CRISPR/Cas9 载体,编辑范围小。SaCas9 长度比 SpCas9 小。
SaCas9 的主要优势是腺相关病毒 (AAV) 包装:AAV 载体将 SpCas9 与 AAV 包装在一起可能具有挑战性。相对较小的 SaCas9 利用 AAV 载体进行 CRISPR 基因编辑成为可能。考虑到这一结构的免疫原性较低,因此 SaCas9 更适合于体内编辑应用,如用于治疗。
CRISPR 2.0
经典的 CRISPR 技术无法与基因组的任意部位结合,有时甚至可能会切割错误的位置。且没有关闭按钮,可能会导致过量切割,产生脱靶效应。对此,科学家们通过一系列研究,筛选出针对 CRISPR-Cas9 的小分子抑制剂 (如 HY-136251, BRD0539 是一种 CRISPR/Cas9 小分子抑制剂,可有效抑制 SpCas9),使得对 CRISPR-Cas9 系统有了更有效的控制。总之,即使有着这样那样的局限性,但经典 CRISPR 系统依然是生物学中非常重要的工具。
被称为 "CRISPR 2.0" 的碱基编辑 (Base Editing)——可以进入到细胞中选定的 DNA 位点,直接将一个碱基转换为另一个碱基,而不会造成 DNA 双链断裂。
CRISPR 2.0 方法并不是要替代传统的 CRISPR 基因编辑技术,而是为治疗某些疾病提供其他可选的方案。
如果目标是要插入或敲除某些 DNA 碱基,CRISPR-Cas9 这种方式还是非常有效的。如果目标仅仅是修复某个点突变,碱基编辑 (CRISPR 2.0) 是一种更安全有效的方式。
岁月悠悠,唯有时间能证明 CRISPR/Cas 系统是不是最合适的基因编辑工具。目前还需要进行大量的实践,同时推进所有这些系统的研究。希望在不久的将来,基因编辑技术能安全地应用到人类疾病治疗、农作物培育等各个方面。

