
mTOR 通路是如何调控自噬的
 2020-12-29
2020-12-29 
 MCE小分子
MCE小分子
mTOR 是细胞生长和代谢的主要调节分子,可促进合成代谢过程,如核糖体的生物发生 (Ribosome biogenesis) 以及蛋白质、核苷酸、脂肪酸和脂质的合成,并抑制分解代谢过程,如自噬。mTOR 信号的失调与许多人类疾病有关,包括糖尿病、神经退行性疾病和癌症。
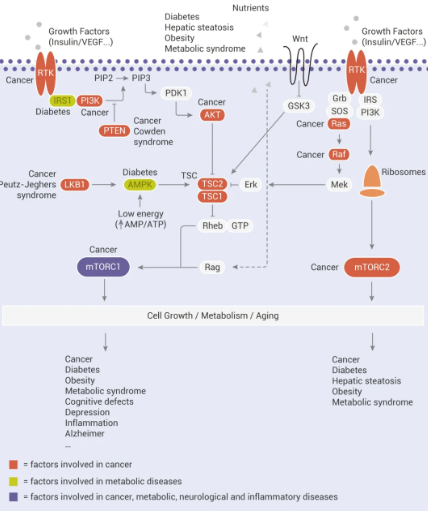
图 1. mTOR 信号通路及对疾病的影响
mTOR 是一种丝氨酸/苏氨酸激酶,属于 PI3K 相关激酶 (PIKK) 家族的成员,可与几种蛋白质相互作用形成两个不同的复合物,分别称为 mTOR 复合物 1 (mTORC1) 和 2 (mTORC2)。mTORC1 包含 mTOR、Raptor、PRAS40、mLST8 和 DEPTOR,对雷帕霉素 (Rapamycin) 敏感;mTORC2 包含 mTOR、Rictor、SIN1、Protor-1、mLST8 和 DEPTOR,对雷帕霉素急性治疗不敏感。
mTOR 通过与 mTORC1 或 mTORC2 的关键组分如 Raptor、Rictor、LST8 和 SIN1 作用,催化多个靶标的磷酸化,如 S6K1、4E-BP1、Akt、PKC、 IGF-IR 和 ULK1,从而调节蛋白合成、营养代谢、生长因子信号传导、细胞生长和迁移以及调节自噬等。

mTORC1 受多种刺激 (例如生长因子、营养物质、能量和应激信号) 以及基本信号传导途径 (例如 PI3K、MAPK 和 AMPK) 激活。
1、生长因子和激素 (例如胰岛素) 通过 I 类 PI3K 及其下游效应子 AKT 的激活来调节 mTORC1 信号传导,从而逆转 TSC1/TSC2 复合物和 PRAS40 对 mTORC1 信号传导的抑制作用;
2、营养物质如氨基酸通过促进 RAS 相关 GTP 结合蛋白 (RAG) 异二聚体向活性构象的转化来刺激 mTORC1;
3、细胞能量状态如低 ATP 水平下,通过 AMPK 磷酸化 TSC2,mTORC1 活性受到抑制。除 PI3K 和 AMPK 途径外,RAS/MAPK 信号也触发了 mTORC1 信号的激活。此外,Hippo 途径 (Hippo 途径与靶向策略)和 Wnt 途径 (Wnt 信号通路很复杂?) 也参与了对 mTORC1 和 mTORC2 的调控。
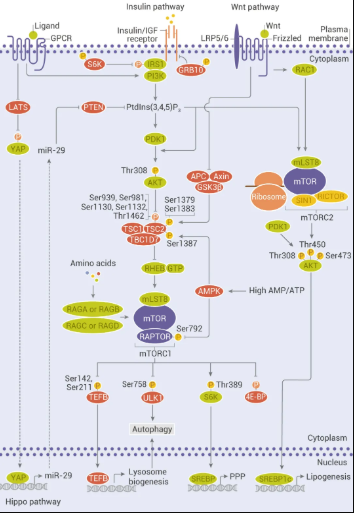
图 2. mTOR 上游:经典和非经典输入
mTORC1 活化后,通过磷酸化下游效应因子 S6K 和 4E-BP 促进合成代谢过程,例如蛋白质、脂质和核苷酸合成,同时,通过 ULK1 抑制分解代谢程序,从而导致自噬抑制。与 mTORC1 相比,对 mTORC2 的了解较少。如 mTORC2 可防止 AKT 泛素化和降解。mTORC2 还可激活 SREBP1c 促进成脂基因表达,以及刺激胰岛素样生长因子 2 (IGF2) 的产生。

自噬是一种溶酶体降解系统,涉及从头自噬形成。而自噬体的形成是由一系列称为 Atg 或“自噬相关”蛋白的蛋白质的协同作用驱动的。成熟的自噬体与溶酶体融合后,会被酸化,形成自溶酶体。溶酶体与自噬体的融合降解捕获的蛋白质,脂质,碳水化合物,核酸和细胞器 (Hello~自噬)。
大量研究表明 mTORC1 是自噬的关键调节剂,调节自噬过程中不同步骤 (如成核,自噬体延伸,自噬体成熟和终止) 。因此,mTORC1 是调控自噬的极具潜力的靶标。
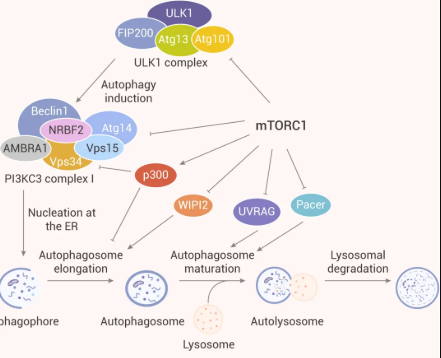
图 3. mTORC1 对自噬各个步骤的调控[9]

1、研究表明,mTORC1 通过磷酸化使形成的自噬调节复合物 (由 ULK1 和其互作蛋白 Atg13、FIP200、Atg101 等形成) 失活,从而影响自噬小体的生物发生。在营养丰富的条件下,mTORC1 通过介导 ULK1 (Ser637 和 Ser757) 和 Atg13 (Ser258) 特定位点磷酸化,抑制 ULK1 复合物的自噬促进激酶 (Autophagy-promoting kinase) 活性。在饥饿和细胞应激期间,mTORC1 活性被抑制,于是与 ULK1 分离。因此,ULK1 和 Atg13 的特定位点的磷酸化被解除。同时,ULK1 复合物通过 Thr180 处自磷酸化而变得活跃,并磷酸化 Atg13、FIP200、Atg101 和其他 Atg 蛋白。活跃的 ULK1 复合物随后转移到内质网的隔离膜上,自噬启动。
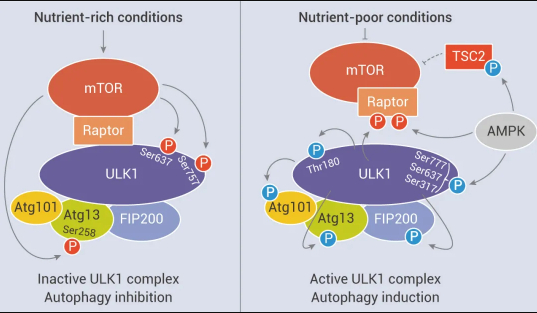
图 4. mTORC1 对 ULK1 复合物的调控作用[9]
2. 另外,AMPK 也与自噬有关。在葡萄糖充足的情况下,活性 mTORC1 通过磷酸化 ULK1 特定位点 (Ser 757) 并破坏 ULK1 与 AMPK 之间的相互作用来阻止 ULK1 活化,抑制自噬启动。在葡萄糖不足的情况下,AMPK 被激活,mTORC1 的磷酸化而被 AMPK 抑制,随后 ULK1 可以与 AMPK 相互作用并被 AMPK 磷酸化,活化的 ULK1 启动自噬。
3、自噬的成核步骤受到 PI3KC3 复合体 I 中 Atg14、AMBRA1 和 NRBF2 的磷酸化抑制。因此,mTORC1 可以通过磷酸化其成分 Atg14,AMBRA1 和 NRBF2 直接调节 PI3KC3-CI 的活性。
4、研究表明,mTORC1 通过分别靶向 WIPI2 和 p300 乙酰转移酶参与调节自噬体形成的延伸步骤,以及 LC3 与自噬体膜的结合。如通过抑制 p300 分子内自抑制,可促进 LC3 的乙酰化,这阻碍了 LC3 的脂化。WIPI2 通过与 Atg16L 结合并将 Atg12-Atg5-Atg16L 复合物结合到吞噬细胞上来促进磷脂酰乙醇胺对 LC3 的脂化作用。
5、mTORC1 还可以通过调节溶酶体生物发生所需基因的转录来间接抑制自噬。如 TFEB 是溶酶体生物发生和自噬基因的主要转录调节因子。TFEB 可上调与自噬体形成、自噬体与溶酶体的融合相关,以及溶酶体生物发生所需的一系列基因。此外,TFEB 的过表达可增加 UVRAG、WIPI、MAPLC3B、SQSTM1、VPS11、VPS19 和 ATG9B 的表达,它们参与了自噬的各个步骤。此外,mTORC2 也可通过激活 mTORC1 间接抑制自噬。
由于自噬与运动、代谢适应和衰老等生理过程,以及神经退行性疾病、传染病、心血管疾病,癌症等疾病过程有关,但自噬诱导是利是弊,取决于疾病的类型或阶段。例如,当营养有限时,自噬可以通过向肿瘤细胞提供营养来促进肿瘤发生和发展。因此,自噬的抑制可能使癌细胞对代谢应激条件敏感,从而导致细胞死亡。另外,mTORC1 作为自噬的主要调节剂,参与自噬过程,是极具有潜力靶标,实际上,mTORC1 的失调已经牵涉到与自噬缺陷相关的疾病中,并且已经有 mTOR 抑制剂在临床试验中或获批用于治疗这些疾病。
总之,mTOR 促进合成代谢并抑制自噬诱导,而用 mTOR 抑制剂调节自噬为多种疾病提供了新的治疗策略。
具有诱导自噬作用的 mTOR 抑制剂
有效且特异性的 mTOR 抑制剂,作用于 HEK293 细胞,抑制 mTOR,IC50 为 0.1 nM;与 FKBP12 结合且抑制 mTORC1;还是一种自噬 (autophagy) 激活剂,免疫抑制剂。
CCl-779
mTOR 抑制剂,IC50 值为 1.76 μM;能激活自噬 (autophagy),在动物模型中防止心脏功能恶化。
WYE-354
ATP 竞争性的 mTOR 抑制剂,IC50 为 5 nM;能抑制 mTORC1 和 mTORC2;在体外能诱导自噬 (autophagy) 激活。
Metformin二甲双胍
抑制肝脏中的线粒体呼吸链,导致 AMPK 活化,增强胰岛素敏感性,可用于 2 型糖尿病的研究;可以透过血脑屏障,诱导自噬 (autophagy)。

