
分子胶改变蛋白-蛋白关系的时空开关
 2020-01-22
2020-01-22 
 MCE小分子
MCE小分子
小分子降解剂是现小分子发展的重要前沿,此前 M 博士已经给大家介绍过一种小分子降解剂——PROTAC,今天我们一起来认识一下 PROTAC 的兄弟 “分子胶” 吧~
小分子抑制剂是靶向治疗的主要类型之一,但也有它的局限性,如无法完全阻断蛋白功能,会引起的脱靶效应和副作用等。然而近年来,小分子降解剂颠覆了先前认为 “不可治疗” 靶点的观点。这些小分子降解剂不仅选择性高,用量小,而且大大降低脱靶效应。
今天我们且来说一说 “小分子降解剂” 的那些事儿~
什么是分子胶?
小分子降解剂通过介导泛素化连接酶与靶蛋白的识别,对靶蛋白进行降解。目前蛋白降解剂主要有两类,一类是双功能分子降解剂,通常称为 PROTAC (蛋白水解靶向嵌合体),通过泛素-蛋白酶体途径特异性地降解靶蛋白 (干货 | PROTAC 知多少)。另一类是 “分子胶” (Molecular glues),也是通过结合泛素化连接酶,但与 PROTAC 不同的是,分子胶是通过修饰泛素化连接酶表面,从而识别并降解全新底物 (Neo-substrate)。例如磺胺类抗肿瘤药 (Indisulam); 和免疫调节药物 (IMiDs) 沙利度胺 (Thalidomide)、来那度胺 (Lenalidomide)。
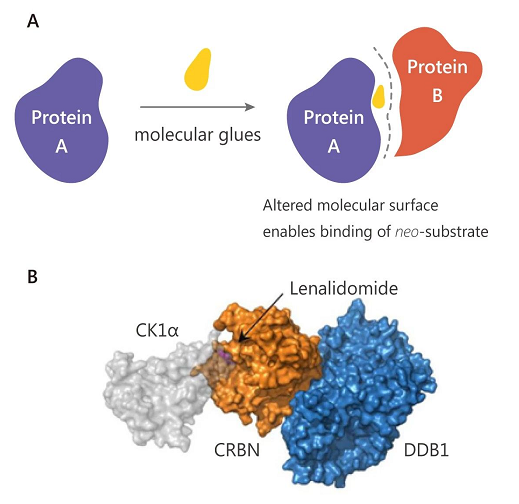
A.“分子胶” 作用原理 B. 来那度胺 (Lenalidomide) 的作用机制
理论来说,PROTAC 分子既可以作为 PROTAC 降解剂来结合和降解靶蛋白,也可以充当 “分子胶” 降解新底物。最近就发现一类邻苯二甲酰亚胺共轭的 PROTAC 降解剂其实是以分子胶的方式起作用,例如 MG-277。另外,最近 Benjamin Cravatt 团队开发出一种新型的“分子胶”。这类“分子胶”是以共价修饰 E3 连接酶 DCAF16 的方式,特异性促进核蛋白降解。
PROTAC “变身” 分子胶
MG-277 是对 MD-222 (PROTAC MDM2 降解剂) 进行了简单的结构修饰而来的。但正是这一小步的修饰,却带来了完全不同的靶向降解 “结局”。
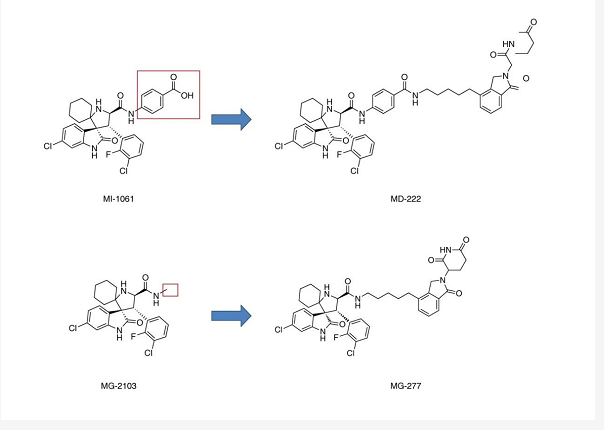
MG-277: MD-222 抑制剂部分的 MI-1061 的苯甲酸被甲基取代
首先,不同于 MD-222 对野生型 p53 (人白细胞 RS4;11) 的选择性,MG-277 对携带野生型,突变 p53 (RS4;11/IRMI-2 细胞) 或缺失 p53 的癌细胞系都有抑制作用,而且在诱导 MDM2 降解方面要比 MD-222 要低得多,且不能激活野生型 p53。通过敲除 MDM2 等试验发现,MG-277 的对细胞的抑制作用不仅不依赖 MDM2,本身也不与 MDM2 结合。
MG-277 对细胞的抑制活性,到底从何而来?
通过对 MG-277 中的戊二酰亚胺的氨基 (与 cereblon 结合相关) 甲基化,即合成了 MC-024,发现 MC-024 在抑制细胞 (RS4;11,RS4;11/IRMI-2,MDA-MB-231 和 MDA-MB-468) 生长方面的效率要低得多。在来那度胺与 MG-277 竞争实验中,来那度胺降低了 MG-277 对上述细胞系的生长抑制。因此,MG-277 对野生型和突变型 p53 细胞生长抑制活性,需要与 cereblon 结合。 据此,作者推测,MG-277 实际上可能起的分子胶的作用,通过促进某些细胞生长相关蛋白 -E3 连接酶 (CRL4CRBN) 复合物募集,进行泛素化和随后的蛋白酶体降解。
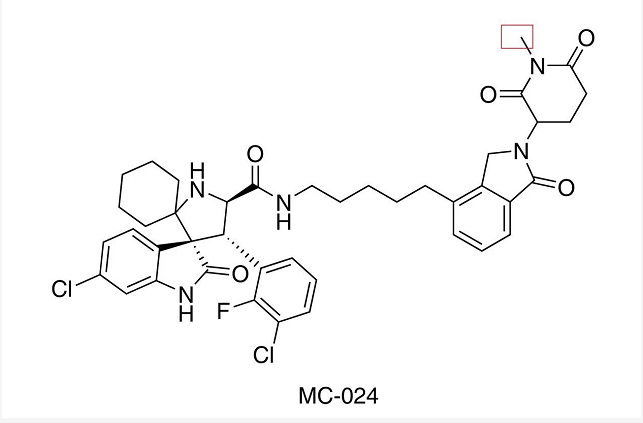
如果 MG-277 作为分子胶,真正的靶标蛋白是什么?
通过对野生型 p53 细胞和携带突变的 p53 细胞蛋白质组分析,MG-277 的靶标蛋白为 GSPT1,且降解 GSPT1 的水平取决于其与 cereblon,蛋白酶体和 Cullin-RING E3 连接酶功能的结合。MG-277 捕获了一些与相互作用有关的关键残基 (cereblon 中 Phe150 和 Glu377),这些残基对于将 GSPT1 募集到 cereblon 中是至关重要的。
总而言之,MG-277 以不依赖 p53 和 MDM2,但以依赖 cereblon,CUL4 E3 泛素连接酶和蛋白酶体的方式,在癌细胞中快速诱导 GSPT1 降解。MG-277 诱导 GSPT1 降解是其细胞生长抑制活性的原因。
而设计的邻苯二甲酰亚胺共轭降解物,既可以作为目的蛋白质的真正降解剂,也可以作为将新底物蛋白募集到 CRL4CRBN 的分子胶,用于泛素化和随后的降解。因此,在 PROTAC 降解剂的设计中需要考虑这两种机制。
特异性降解 “核蛋白” 的分子胶
接着,我们来讲讲另外一种新型“分子胶”。Benjamin Cravatt 团队找到了新的 E3 连接酶成员 DCAF16,一种目前没有文献报道过相关功能的多肽 (在哺乳动物中高度保守,但啮齿动物却不存在的多肽),此类 “分子胶” 以共价修饰 DCAF16 的方式,特异性促进核蛋白降解。
DCAF16连接酶的发现
Benjamin Cravatt 团队在之前的工作中鉴定出一些与半胱氨酸结合共价片段,作者称为 “侦探” 片段 (Scoutfragment)。它们能 “搜寻” 出人类蛋白组中可配体的半胱氨酸,包括一些从未被研究的泛素化连接酶。作者将 KB02、KB03、KB05 三个“侦探”片段连接在 FKBP12 高亲和性的配体 SLF 上。发现 KB02-SLF、KB03-SLF、KB05-SLF 均未改变胞质蛋白 FKBP12 水平。其中,KB02-SLF 显著降低核蛋白 FKBP12 的水平。
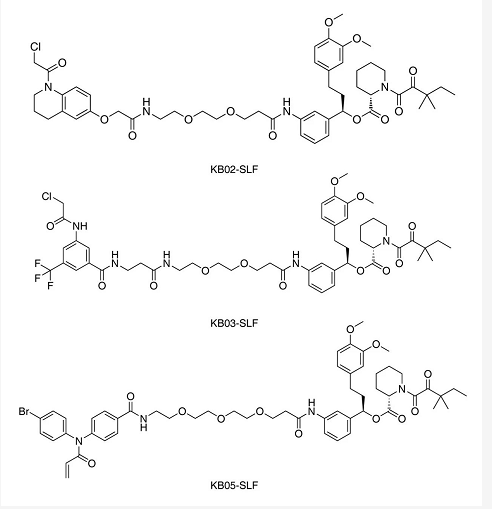
选择性的 Cullin-RING 泛素连接酶 (CRL) 类 E3 连接酶抑制剂 MLN4924 阻断了 KB02-SLF 介导的核 FKBP12 降解,表明 KB02-SLF 是通过 CRL 的作用促进了核定位 FKBP12 的蛋白酶体降解。通过蛋白质组学以及 DCAF16 的 shRNA 沉默进一步确定 CRL 类 E3 泛素化连接酶为 DCAF16。
DCAF16在诱导核蛋白降解的过程中是如何被修饰的?
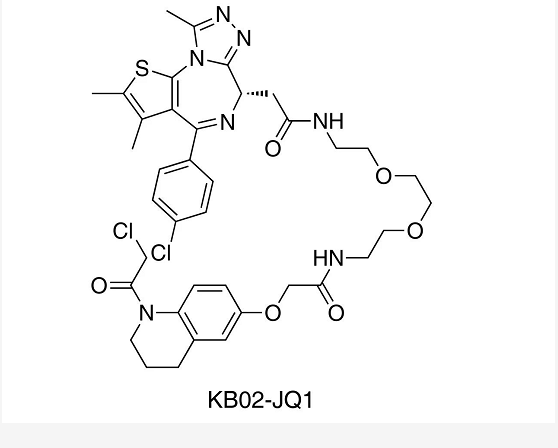
串联质谱 (MS/MS) 分析和定点诱变实验表明,KB02-SLF 是修饰了 DCAF16 的 Cys177-179 三个半胱氨酸位点。其中 DCAF16 中的 C177 和 C179 为极有可能是介导 FKBP12 降解的关键的位点。接下来,作者构建的 KB02-JQ1 双功能分子 (诱导核蛋白 BRD4 降解),在敲除的 DCAF16 细胞中,KB02-JQ1 诱导的 BRD4 降解则被抑制。
DCAF16 的发现,在 CRBN 和 VHL 的蛋白降解剂的基础上,拓展了蛋白降解剂的开发与应用前景。同时,特异性促进核蛋白的降解,避免降解了同双功能化合物结合的胞质蛋白。另外,较少成分的 DCAF16 修饰就可支持配体诱导的蛋白质降解,而未被修饰的 DCAF16 仍然能调节其底物,使蛋白降解剂能最小化受到 DCAF16 内源底物的干扰。
相关产品的文章推荐
MG-277
MG-277 是一种由 PROTAC 降解剂转化而来的分子胶。MG-277 诱导中等程度的 MDM2 降解。MG-277 以非 p53 的方式高度有效地抑制肿瘤细胞的生长,对 RS4;11 细胞和 p53 突变体 RS4;11/IRMI-2 细胞的 IC50 分别为 3.5 nM 和 3.4 nM。MG-277 有效的诱导翻译终止因子 GSPT1 的降解,DC50 为 1.3 nM。MG-277 具有有效的抗癌活性。
KB02-SLF
KB02-SLF 是一种核 FKBP12 降解剂 (molecular glue)。KB02-SLF 通过共价修饰 DCAF16 (E3 ligase) 促进 FKBP12 降解,并可以提高生物系统中蛋白质降解的持久性。KB02-SLF 由 SLF 与泛素 E3 连接酶配体 (KB02) 通过 linker 连接而成。
KB02-JQ1
KB02-JQ1 是一种高选择性 BRD4 降解剂 (molecular glue)。KB02-JQ1 通过共价修饰 DCAF16 (E3 ligase) 促进 BRD4 降解,并可以提高生物系统中蛋白质降解的持久性。KB02-JQ1 由 JQ1 与泛素E3 连接酶配体 (KB02) 通过 linker 连接而成。
Pevonedistat
Pevonedistat (MLN4924) 是一种有效,选择性的 NEDD8 活化酶 (NAE) 抑制剂,IC50 为 4.7 nM。NEDD8 活化酶参与 CRL 激活的第一步,通过激活 NEDD8 并随后转移到 Ubc12,NEDD8 途径的 E2 结合酶。

