
MCE解析5mC甲基化之路
 2019-12-20
2019-12-20 
 MCE小分子
MCE小分子
什么是“甲基化”,DNA 为什么会发生“甲基化”,又是怎么发生的?。。。说起 "DNA 甲基化”萌 CeCe 是满脑子问号啊,于是又抱住小 M 博士的大腿求开讲了~
表观遗传学被定义为“不依赖于 DNA 序列的变化而是由于染色体改变所产生的稳定可遗传的表型”。DNA 甲基化,组蛋白结构的改变以及 microRNA 对基因的调控等都是表观遗传信息的重要载体。其中 DNA 甲基化作为重要的表观遗传修饰之一,对基因组稳定性,转录和发育产生深远影响。
甲基化到底是什么?
胞嘧啶第五碳的甲基化 (5- 甲基胞嘧啶:5mC) 是最早在真核生物中被挖掘出的甲基化类型,它也是今天我们要介绍给大家的重点。早在几十年前,就有体内外研究表明,它与转录抑制有关。
哺乳动物中,5mC 往往存在于 DNA 双螺旋中的鸟嘌呤 (G) 的上游,即所谓的 CpG 位点。除了 5mC,其他的甲基化类型有:4- 甲基胞嘧啶 (4mC),5- 甲基胞嘧啶 (5mC) 和 6- 甲基腺嘌呤 (6mA)。
DNA 甲基化具有多种功能:与转座子和基因的抑制有关,但也与活跃转录的基因体 (gene body) 有关,在某些情况下还与基因激活本身有关。
DNA 是如何出现表观遗传修饰或标记呢?下面以哺乳动物中 5mC 为例,揭开 DNA 甲基化修饰的神秘面纱!
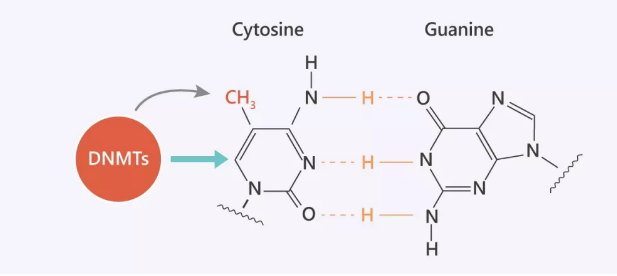
图 1. DNA 复制过程中 CG 配对
Tip 1: CpG 是胞嘧啶 “C” 和鸟嘌呤 “G” 的缩写,由磷酸酯 “p” 连接。在哺乳动物中,胞嘧啶甲基化主要发生在 CpG 二核苷酸中。可通过对二核苷酸序列 5'CpG3' 中胞嘧啶碱基的甲基化共价修饰脊椎动物的 DNA。
DNA 甲基化分为三个阶段:甲基化的建立 (从头开始 DNA 甲基化),甲基化的维持和脱甲基。甲基化从写入到擦除,牵涉着 DNMT 与 TET 两大蛋白家族的“爱恨情仇”。
增添 DNA 甲基化—— DNMT 建立并维持
DNA 甲基转移酶 (DNMTs) 是 DNA 甲基化的写入蛋白。DNMT1 使用 S- 腺苷甲硫氨酸 (SAM) 作为甲基供体,将甲基“搬到” DNA 上 (如图 Fig 1)。

图 2. 5mC 甲基化
DNMTs 家族蛋白又可细分为从头甲基转移酶 (DNMT3A, DNMT3B 和 DNMT3L) 和维持甲基转移酶 (DNMT1) 两大派别,在甲基化中发挥不同的作用。
从头甲基化:(De novo of DNA methylation )
DNA 甲基化通过称为 DNMT3A 和 DNMT3B “从头甲基化酶”在胚胎发育过程中建立。DNMT3A 和 DNMT3B 是对未甲基化的 DNA 具有活性的从头甲基转移酶,他们就像是甲基化历程的“创业者”,不偏好半甲基化的 CpG 位点,“从头”甲基化先前未被甲基化的 DNA。
DNMT3 家族还存在一个成员:DNMT3L。与 DNMT3A/3B 相比,缺乏甲基基序和催化结构域,但可以与 DNMT3A/3B 结合,刺激 DNMT3A/3B 的酶活性,在甲基化进程中推波助澜。
维持甲基化:(Maintenance of DNA methylation)
维持甲基化,就是双链 DNA 的一条链已经发生甲基化,已经发生甲基化的链想将这种甲基化状态“薪火相传”(通过半保留复制)。为完成这一步骤,“维持甲基转移酶” (DNMT1) 上线,DNMT1 更像是个猎头,只认识甲基化 CpG,没有甲基化的 CpGs 不会成为维持甲基转移酶 DMNT1 的底物。在这个进程中,DNMT1 相互作用蛋白 UHRF1 (多结构域蛋白 E3 泛素蛋白连接酶) 至关重要,因为 UHRF1 的 SET 和 RING 相关 (SRA) 域对半甲基化的 DNA 具有强烈的偏好,能帮助 DNMT1 在复制叉处靶向底物 CpG 二核苷酸。
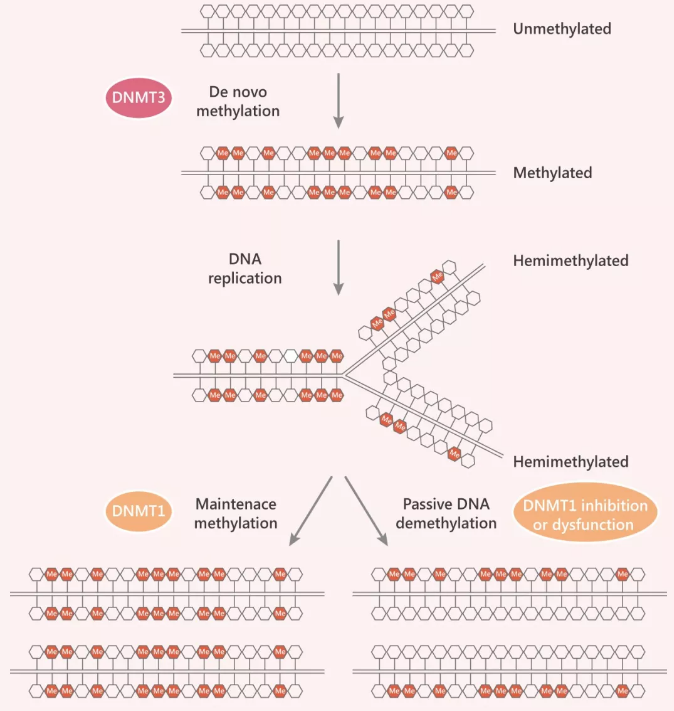
图 3. DNA 的从头甲基化与维持甲基化
Tip 3:为确保亲代模式在子代链上的复制,一种“维持甲基转移酶” DNMT1,只会将甲基化 CpGs 碱基与甲基化的亲代 CpG 配对,非甲基化 CpGs 不会成为其底物。这种简单机制的结果是,DNA 甲基化模式会像 DNA 自身的碱基序列一样被半复制。
除 DNA 甲基化——擦除蛋白 -Tet 启动的 DNA 脱甲基途径
DNA 脱甲基涉及被动和主动脱甲基的两个过程来实现:
5- 甲基胞嘧啶 (5mC) 作为 DNA 修饰的主要形式。结构上,甲基通过牢靠的碳-碳双键与胞嘧啶碱基的 5 位连接,为直接去除甲基加了一层保险。机制上,前文提到的 DNMT3 以及官能伙伴 UHRF1 一起协助维持这种状态。尽管如此,5mC 依然有自己的办法逃脱层层枷锁。
被动 DNA 脱甲基化:
功能性的 DNA 甲基化维持机制缺乏,使得 DNA 复制过程中 5mC 的逐步‘稀释’ (passive solution),这个过程通常被称为被动 DNA 脱甲基化 (参照 Fig 2)。该事件主要发生在分裂细胞中,DNMT1 的抑制或功能障碍会使新掺入的胞嘧啶保持未甲基化,润物细无声地降低了每个细胞分裂后的总体甲基化水平。
主动 DNA 脱甲基化:
敲黑板喽,该脱甲基化的过程,也称为活性去甲基化。在这个过程中 TET 蛋白家族起到了举足轻重的作用 (参照 Fig 3)。
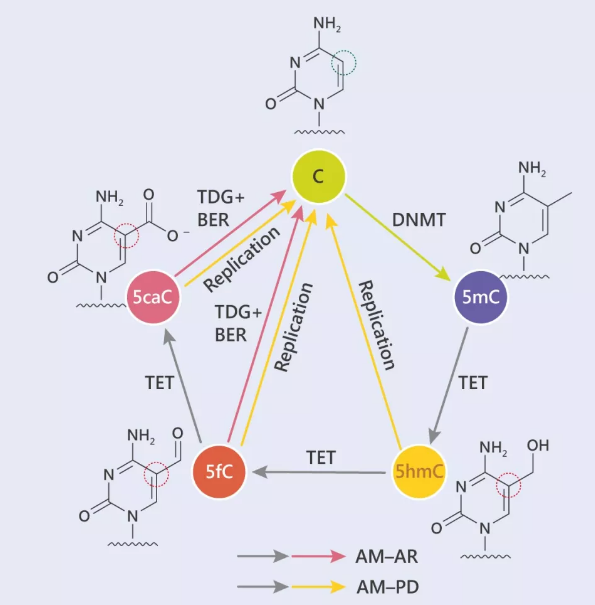
图 4. Tet 启动的 DNA 脱甲基 (氧化位点已圈出)
在主动脱甲基化的过程中,5mC 通过 TET 蛋白,不断被氧化,一步步去除“甲基化帽子”。
这个过程中。TET 蛋白像个探测器一样,可以沿着 DNA “拉链”从一个点滑动到另一个 CpG 位点,最神奇的是,TET 会优先识别并且氧化同一 DNA 分子上的其他 CpG 位点。
TET 蛋白可以介导 5mC 迭代氧化为 5- 羟甲基胞嘧啶 (5hmC),5- 甲酰基胞嘧啶 (5fC) 和 5- 羧胞嘧啶 (5caC)。氧化的 5mC 衍生物不能用作 DNMT1 的底物,在复制的过程中被动脱甲基作用而丢失,胸腺嘧啶DNA糖基化酶 (TDG) 介导的 5fC 和 5caC 切除的复制依赖稀释,再加上碱基切除修复 (BER),导致去甲基化,这个过程被定义为主动修饰-主动去除 (AM-AR)。此时的 5fC 和 5caC,就像是无业游民,TDG 能识别这种不正常的碱基并切除它们,随后,AP 位点形成 (无嘌呤或者无嘧啶位点),开始碱基切除修复。另外,也可以通复制依赖 5hmC,5fC 或 5caC 的方式稀释 DNA 甲基化,即主动-被动去除 (AM-PD)。
Tip 4: TDG :能够识别不正常的碱基。如胞嘧啶 5mC 在脱氨基之后会直接形成尿嘧啶 5hmc,对于这些不正常的尿嘧啶,就可以被 TDG 切除掉,形成无嘌呤或者无嘧啶的位点。Base-excision repair:AP 位点形成,AP 核酸内切酶在 AP 位点将 DNA 链切开,然后核酸外切酶 1 将包括 AP 位点在内的 DNA 链切除。这种单个碱基的修复叫做碱基切除修复。
识别 DNA 甲基化——读取蛋白
如果将甲基化的 DNA 比作是一匹千里马的话,那么三个识别的蛋白家族 (MBD 蛋白质,UHRF 蛋白质和锌指蛋白) 算得上是识别它的伯乐了。
MBD 家族由 MeCP2 和 MBD1-6 组成,共享一个保守的 MBD 域,这也与甲基化 DNA 结合所必需的。MeCP2,MBD1 和 MBD2 还包含一个 TRD 域,可帮助他们募集染色质重塑核心加压因子 (chromatin remodeling corepressors),从而引起转录沉默。如 MeCP2 的 TRD 结构通过募集包含 Sin3A 共阻遏物和组蛋白脱乙酰基酶 (HDAC1 和 HDAC2) 的染色质重塑复合体来参与介导基因沉默。
UHRF 蛋白 (如 UHRF1 和 UHRF2 ),前面在维持甲基化中已经提过它的作用了。锌指蛋白 (Kaiso,ZBTB4 和 ZBTB38) 则是通过锌指结构域与甲基化 DNA 结合。其中 Kaiso 被认为是转录阻遏物,可以与活跃表达基因的未甲基化区域结合。
DNA 甲基化与组蛋白修饰
DNA 甲基化和组蛋白修饰的关系千丝万缕。简而言之,DNA 甲基化有助于引导组蛋白修饰,组蛋白修饰又会指导 DNA 甲基化。例如 DNMT3A 的 PWWP 域会与 H3K36me3 相互作用,增强 DNMT3A 的活性。同样,组蛋白修饰在 DNA 甲基化机制中也起着重要作用。拿 DNMT1 启动子过程中的甲基化来说,H3K4me3 可抑制DNMT3 酶。而在没有 H3K4 甲基化的情况下,抑制会得以解除,从而使 DNA 甲基化。
H3K4me3 标记了活跃的启动子,阻止了 DNMT3A 和 DNMT3B (以及DNMT3L) 的 ADD 域的染色质结合,从而使 ADD 与甲基转移酶 (MTase) 结构域结合并自动抑制 DNMT3 酶。在没有 H3K4 甲基化的情况下,ADD 域与 H3K4 结合,并且自动抑制得以解除,从而 MTase 域可以使 DNA 甲基化。
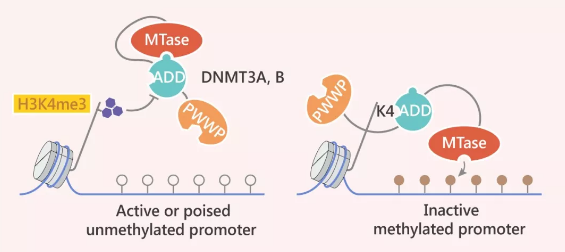
图 5. 启动子 DNA 甲基化的机制
Tip 5:H3K4me3 标记了活跃的启动子,阻止了 DNMT3A 和 DNMT3B (以及DNMT3L) 的ADD域的染色质结合,从而使ADD 与甲基转移酶 (MTase)结构域结合并自动抑制 DNMT3 酶。在没有H3K4甲基化的情况下,ADD 域与 H3K4 结合,并且自动抑制得以解除,从而 MTase 域可以使 DNA 甲基化。
癌症中的 DNA 甲基化
在高等动物中,癌细胞中的 DNA 甲基化模式明显“扭曲”。大约 70% 的基因拥有与启动子相关的 CpG 岛 (CG 碱基频率高的区域),而维持未甲基化的启动子 CpG 岛对提高转录潜能有积极作用,并与活性的组蛋白修饰如组蛋白 H3 和 H4 的乙酰化和 H3K4 的甲基化有关。
相反,癌细胞明显表现出 CpG 岛的密集高甲基化,包括基因间区和重复元件在内的大量染色质的低甲基化。密集甲基化的 CpG 岛能够让原本松散的染色质紧缩在一起,抑制与“抑制性修饰” (H3K9me2/3,H3K27me3 和/或 H4K20me3) 相关的基因表达,可见 DNA 甲基化显著着影响癌细胞的基因组格局。
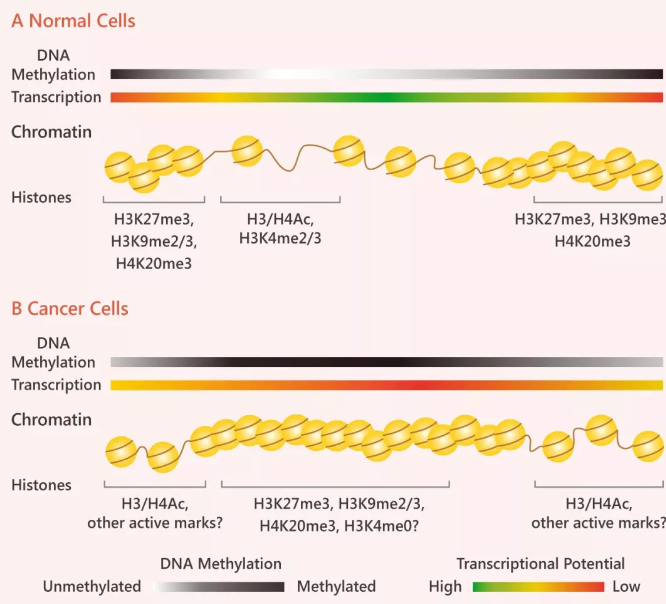
图 6. DNA 甲基化和组蛋白修饰模式在癌症中的改变
Tip 6: DNA 被包裹在组蛋白的八聚体中,形成核小体,即染色质的最小单位。
正常情况下转录时是开放松散的染色质结构:组蛋白 H3 和 H4 的高度乙酰化,组氨酸 H3 在赖氨酸 4 (H3K4me2/2/3) 处的二甲基和三甲基化 (Fig 5A)。然而,当转录由于被甲基化受阻时,受阻区域染色质结构就会变得紧缩在一起,H3/H4 乙酰化和 H3K4 甲基化不显示,取而代之的是富含 H3K9 (H3K9me2/3),H3K27的三甲基化 (H3K27me3) 和 H4K20 的三甲基化 (H4K20me3)(Fig 5)。
DNA 甲基化抑制剂
DNA 甲基化抑制剂一般分为核苷类似物和非核苷类似物化合物。核苷类似物如 Decitabine,5-Azacytidine,Zebularine,它们可掺入 DNA 中,掺入后,它们充当 DNMT 酶的自杀底物。非核苷化合物可以抑制 DNA 甲基转移酶活性,如 RG108,DC-05,它们直接阻断 DNA 甲基转移酶的活性。
Decitabine,地西他滨,FDA 批准的 DNA 甲基转移酶 (DNMTs) 抑制剂。通常用于治疗骨髓增生异常综合征 (MDS) 和急性髓性白血病 (AML)。Decitabine 具有双重的剂量依赖性作用机制。高剂量时,Decitabine 通过掺入 DNA 代替胞嘧啶后,可将 DNA 甲基转移酶共价捕获到 DNA 中,从而诱导细胞毒性作用。在较低剂量下其抗肿瘤效果可能是由于它能够抑制 DNA 甲基化和重新激活肿瘤抑制基因。
5-Azacytidine,阿扎胞苷,DNA 和 RNA 核苷胞苷类似物,也是FDA 批准的 DNA 甲基转移酶 (DNMTs) 抑制剂,5-Azacytidine 必须与 DNA 结合以共价捕获 DNA 甲基转移酶。5-Azacytidine 也可以掺入到 RNA 中。
RG108,非核苷类似物,是一种靶向人 DNA 甲基转移酶 (DNMTs) 的抑制剂,
IC50 值为 115 nM。RG108 结合并阻断 DNMTs 的活性位点。RG108 缺少与 Decitabine 相关的高水平的细胞毒性。RG108 可在不影响着丝粒卫星序列甲基化状态的情况下,引起肿瘤抑制基因的去甲基化和再激活。

