
内质网应激库,MCE 独家上线
 2020-04-17
2020-04-17 
 MCE小分子
MCE小分子
上周,小编带大家了解了下,内质网应激中,PERK、IRE1、和 ATF6 三种重要的信号转导通路,感应并传导信号,"指示" 一系列保护反应如 UPR。那么,那些“不合格”蛋白后来怎么样了呢?"垃圾" 当然要分类处理,不能 "活着" 出内质网。
AvadaKedavra (阿瓦达索命) 是哈迷们灰常熟知的死亡咒语,今天看完内质网 (ER) 的 "阿瓦达索命",你会发现科学界的 "魔法",如此错综复杂,但又如此井然有序。
真核细胞中,ER 是近三分之一蛋白的合成位点,如此重要的结构,必须具备完好有序的机制,才能确保部署到远端分泌途径的蛋白正确折叠加工、运出 ER、正确发挥作用。而 "不合格" 的蛋白会激发降解程序,常见的有内质网相关蛋白降解途径 (ERAD) 和自噬途径。ERAD 是一个复杂的过程,"不合格" 蛋白通过这个过程被筛选,最终被泛素-蛋白酶体系统 (UPS) 降解。ERAD 通过紧密衔接的四个步骤进行,包括底物选择、ER 膜上的转位、多聚泛素的共价结合和蛋白酶体降解。
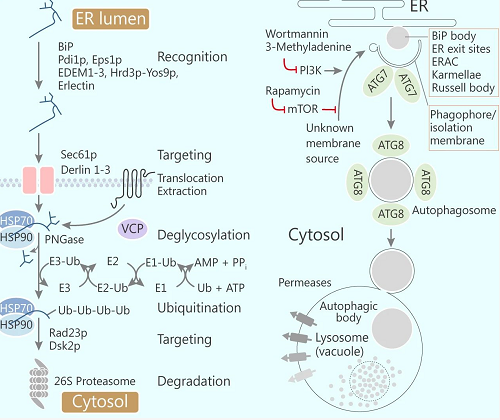
ERAD 和自噬
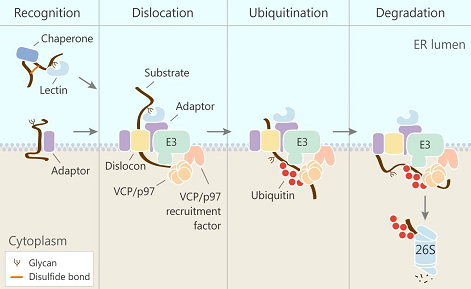
ERAD 的关键四个步骤
Step1: ERAD 识别、选择底物
以糖基化蛋白为例,寡糖转移酶识别大多数新生肽链中的 Asn-X-Ser/Thr 序列,并与 ER 膜上的甘露糖核心糖基 Glc3Man9GlcNAc2 共价结合到蛋白。如果蛋白正确折叠,它可以离开 ER 到达最终目的地。未折叠的蛋白会被保留在 ER 中,并被 UGGT (UDP-葡萄糖-糖蛋白葡萄糖基转移酶)再糖基化,然后在 CNX-CLR (凝集素伴侣分子) 循环中,进一步尝试折叠。但该机制目前尚不清楚。"折叠" 失败的蛋白,脱糖基后可被 EDEM (ER 降解增强子 α-甘露糖苷酶样蛋白) 识别。ER 常驻蛋白 OS-9 和 XTP3-B/Erlectin 通过甘露糖-6-磷酸受体同源 (mannose-6-phosphate receptorhomology, MRH) 结构域选择性地识别 ERManI/EDEM1-3 产生的剪切寡聚糖。最终,EDEM 将通过促进 ERAD 凝集素 OS9 和 XTP3-B 的结合而靶向错误折叠的糖蛋白启动降解程序。
ERAD 的底物识别步骤较为复杂,涉及多种酶和伴侣蛋白的协同作用。例如在退出 CNX-CLR 循环后,EDEM1 进一步修饰了 Man8-GlcNAc2 的糖链结构,ERdj5 减少了二硫键,并激活 BiP 的 ATP 酶活性。BiP·ADP 与错误折叠的蛋白结合,等待促使其转位。
非糖基化蛋白的 ERAD,独立于凝集素样蛋白途径。如免疫球蛋白轻链 (IG-K-LC) 是一种非糖基化的 ERAD 底物,以 BiP 途径降解。OS9 和 XTP3-B 也参与了非糖基化蛋白的 ERAD。
Step2: ER 膜转位
ERAD 底物需要完全或部分 (膜整合蛋白) 进行 ER 跨膜,在胞质中降解。哺乳动物中,转位复合物通过 Sec61p 通道转位。该过程涉及各种适配器,SEL1L 就是一个典型。SEL1L 将管腔底物识别因子与 Hrd1 连接起来。SEL1L 与 OS-9 和 XTP3-B 结合,EDEM1 和 EDEM3 结合,ERFAD-ERp90 结合。SEL1L 与膜 ERAD 组分形成复合物,这些复合物激活 VCP/p97 复合物。因此 SEL1L 是 ERAD 底物招募、转位和泛素化的重要连接。二硫键有助于稳定三级结构,相反的,错误折叠蛋白的二硫键的减少有助于 ERAD。ERFAD (ER 与降解相关的黄素蛋白) 和蛋白二硫键异构酶 (PDI) 家族成员 ERp90/TXD16 与 Hrd1 适配器 SEL1L 结合。ERFAD-ERp90 复合物可以减少 SEL1L 相关的糖蛋白底物的二硫键。ERdj5 (ER 氧化还原酶) 也有类似的活性。
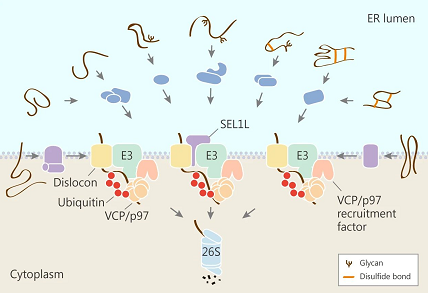
ER 膜内的蛋白适配器将底物识别链接到转位装置
Step3-4: 泛素化和降解
泛素可以作为单体或链与其他蛋白共价结合,通过一个复杂的、高度调控的过程被称为泛素化。泛素链一般附着在错误折叠蛋白的 Lys 残基上。泛素化最确定的作用是通过 26S 蛋白酶体进行蛋白降解。这种靶向性需要通过 Lys48 (K48) 修饰,并通过 26S 蛋白酶体的 19S 帽进行特异性识别。
泛素化包含三种酶: E1 激活酶、E2 偶联酶和 E3 连接酶。泛素化过程中,E1 以三磷酸腺苷 (ATP) 依赖的方式在其活性半胱氨酸和甘氨酸 C 端之间形成巯基酯键。泛素在 E1 上被激活,并通过反式硫代反应转移到 E2 的活性半胱氨酸。E3 既与 E2 结合,又与底物结合,促进泛素的 C 端甘氨酸与底物上的赖氨酸残基形成异肽键。
虽然 Hrd1p 和 Doa10p 在进化上是保守的 (哺乳动物中分别为 Hrd1 和 TEB4),但哺乳动物中 ERADE 3s 的数量却在不断增加。gp78、RNF5/RMA1、RNF170、RNF185、Trc8、RNF103、RFP2、Fbx2、Fbx6、Parkin、CHIP 和 UBE4a 均是。诸多 ERAD 辅助因子也被证明与 Hrd1 相互作用,包括与 VCP/p97 相互作用的 UBXD2 和UBXD8,以及伴侣蛋白 ubiquilin 和 BAG6。
在初始的 E3 介导的泛素连接后,泛素链延长 ("多泛素化") 通过额外的泛素单体对之前连接的泛素的 Lys 残基进行共价修饰而发生。这形成了一个延伸的异肽连接多泛素链。有助于 ERAD 的发生。
胞质中 AAA+ ATPase p97 (酵母中为 VCP 或 Cdc48p) 作为一种重要的蛋白,在多种生物过程中发挥多种作用,如 ERAD、转录调控、自噬、调节外线粒体膜蛋白降解等。VCP/p97 功能的多样性是由其与 N 端相互作用的蛋白的多样性决定的。VCP/p97 与 Hrd1、gp78 等 E3 酶作用,与 ataxin3、YOD1 等 DUBs 作用,与 UbxD2、VIMP 等 ERAD 辅助因子作用 。VCP/p97 作为一种分离酶,利用 ATP 水解的能量将 ER 复合物中的泛素化蛋白分离到细胞质中。已发现至少有七个不同的 ERAD 成员通过特定 "motif" 与 VCP/p97 相互作用,如 VIM motif (gp78 和 SVIP), UBX 结构域 (UBXD2 和 UBXD8),SHP box (Derlin1 和 Derlin2),还有 Hrd1 和 VIMP 未被定义的区域。VCP/p97 能作为支架将转位的底物与参与底物修饰和加工的细胞质辅因子连接起来。胞质去糖基酶 NGly1 通过其 PUB 结构域直接与 VCP/p97 结合而被募集到转位复合物中,其功能是通过从脱位的底物中去除连接的聚糖来促进蛋白酶体对底物的清除。VCP/p97 还与许多 Ub 结合蛋白 (Ufd1 和 Npl4)、去泛素酶 (Yod1、VCIP135、Usp19 和 Ataxin-3) 和 E4 泛素延伸酶 (Ube4a) 有关。
尽管,哺乳动物 ERAD 机制的核心成分的研究已经取得了相当大的进展,但仍有许多问题没有得到解答。如此庞大、复杂但又足够严谨的调控网络,还需要更多的发现和完善。
内质网应激引发的自噬
在自噬过程中,吞噬团或隔离膜吞噬细胞质和细胞器,比如线粒体、过氧化物酶体、部分内质网或外来颗粒,然后与溶酶体或者液泡融合,降解掉这些吞噬物。自噬导致的选择性底物降解需要 ER 的隔离化,吞噬小泡可识别这些隔区。分泌途径中,错误折叠蛋白的累积会促进 ER 隔区和 ER-Goligi 中间小室 (ERGIC) 的增加。这些小室含有大量的错误折叠蛋白和伴侣分子,如 BiP。凝集素伴侣蛋白或可溶性 ER 常驻蛋白的选择性富集,或 ER 常驻蛋白的不可及性预示了 ER 的分室化和分选机制的存在。
番外: ERS 引起的细胞死亡!
当 ERS 持续发生,超出负荷,会引发细胞层面的死亡,涉及的靶点主要有 caspase 家族、DR5/TRAL-R2、RIPK、Atg5 等等,不赘述,看图就好。

持续的 ERS 引起细胞死亡
产品推荐
ISRIB (trans-isomer)
透过 BBB,高效的 PERK 抑制剂;增加长期记忆
IRE1α RNase 激酶抑制剂;抗纤维化活性
Kira8
IRE1α RNase 激酶抑制剂;用于糖尿病研究
ATF-6 激活剂
新型选择性的、ATF-6 抑制剂
Benzenebutyric acid
ERS 抑制剂;癌症和感染等研究

